AppsR+HawkesR-2009¶
Notes about [AppsR+HawkesR-2009] 1.
- 1
Richard Apps and Richard Hawkes. Cerebellar cortical organization: a one-map hypothesis. Nature Reviews Neuroscience, 10:670–681, 9 2009. doi:10.1038/nrn2698, Notes: AppsR+HawkesR-2009.html (this file).
This paper describes different zones of the cerebellum.
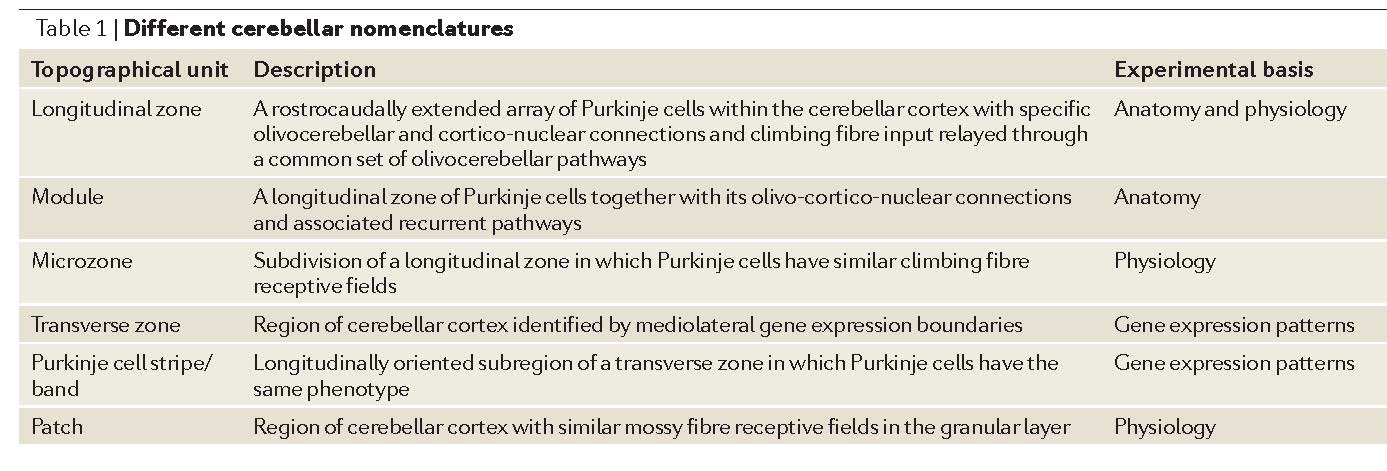
Table 1 in AppsR+HawkesR-2009 [AppsR+HawkesR-2009].¶
671
Transverse zones Although anatomical, physiological and behavioural studies have emphasised a longitudinal organization within the cerebellar cortex (see later), developmental studies17,18 suggest that the development of the fundamental cerebellar architecture begins with the subdivision of the cerebellar cortex into five (or six in birds) transverse zones17 (FIG. 2)
Stripes Every transverse zone is subdivided into a series of stripes (or ‘bands’) oriented along the rostrocaudal axis and which are defined by the restricted expression of molecular markers (FIG. 2). The most comprehensively studied molecular marker is zebrin II22, which cloning studies revealed to be the metabolic enzyme aldolase C23,24. Zebrin II is expressed by a subset of Purkinje cells (zebrin II+) that alternate with Purkinje cells that do not express this marker (zebrin II–), thus forming zebrin II+/– stripes (FIG. 2). The zebrin II+/– stripes are symmetrically distributed across the midline, highly reproducible between individuals22,25,26 and conserved across species (reviewed in REF. 27).
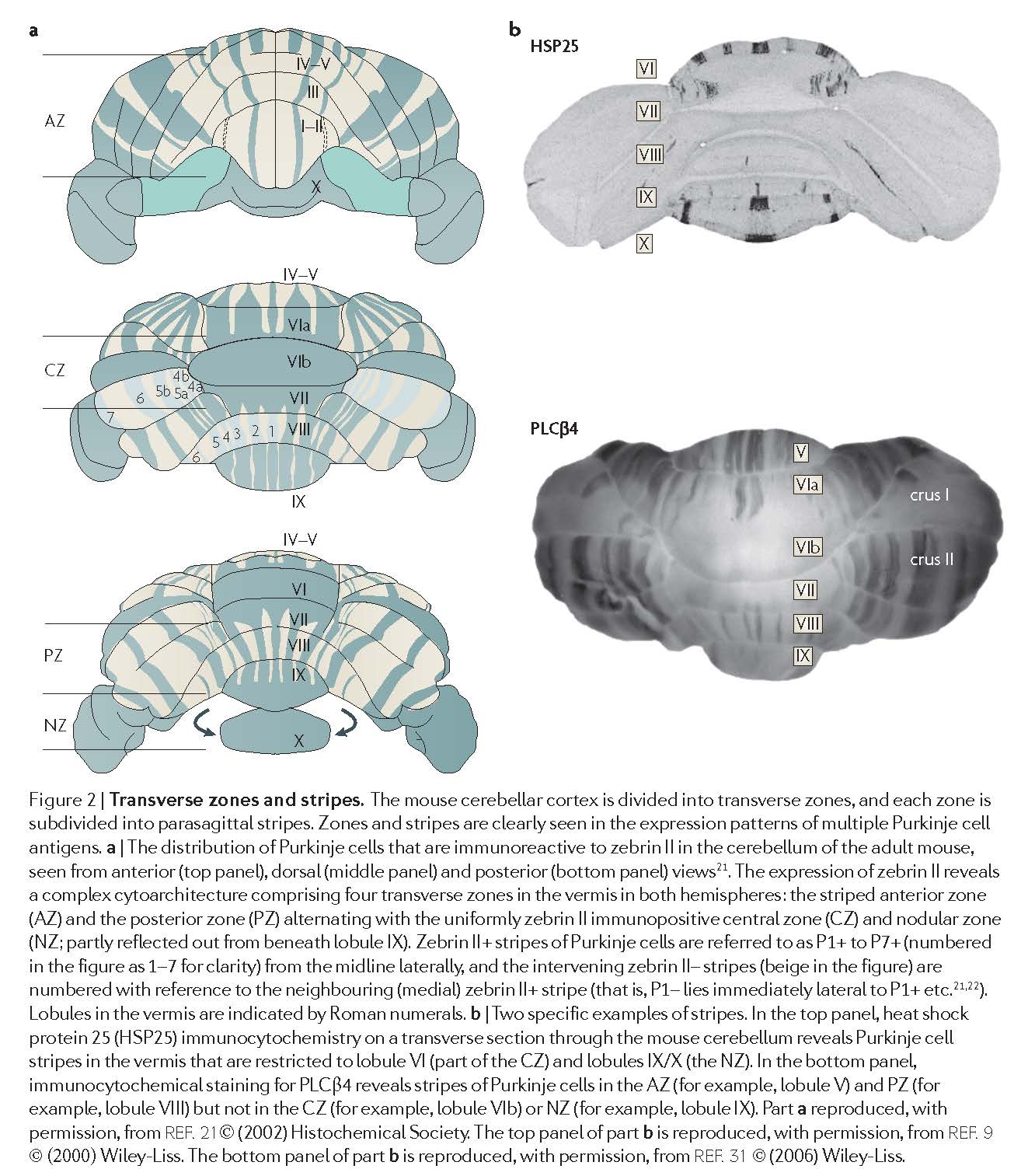
Fig 2 in AppsR+HawkesR-2009 [AppsR+HawkesR-2009].¶
673:
Longitudinal zones Numerous pathway tracing studies have revealed ‘longitudinal zones’ within the cerebellar cortex: narrow, rostrocaudally elongated regions that run perpendicular to the long axis of the lobules3,78–80 (FIG. 4). Longitudinal zones were originally defined by their topographically organized Purkinje cell output to different territories within the cerebellar and vestibular nuclei (the socalled A, b, C1, C2, C3, D1 and D2 longitudinal zones of Voogd; for a review of the earlier literature see REF. 2); additional longitudinal zones have subsequently been added, for example, X, CX and D0 (FIG. 4). A longitudinal zonal arrangement also holds true for the anatomy of olivocerebellar climbing fibre projections81 and for the pattern of termination of physiologically characterised spino-olivocerebellar pathways (as described originally by Oscarsson and co-workers82). In general, the maps of olivocerebellar connections defined by anatomical and physiological methods coincide, for example, see REF. 83.
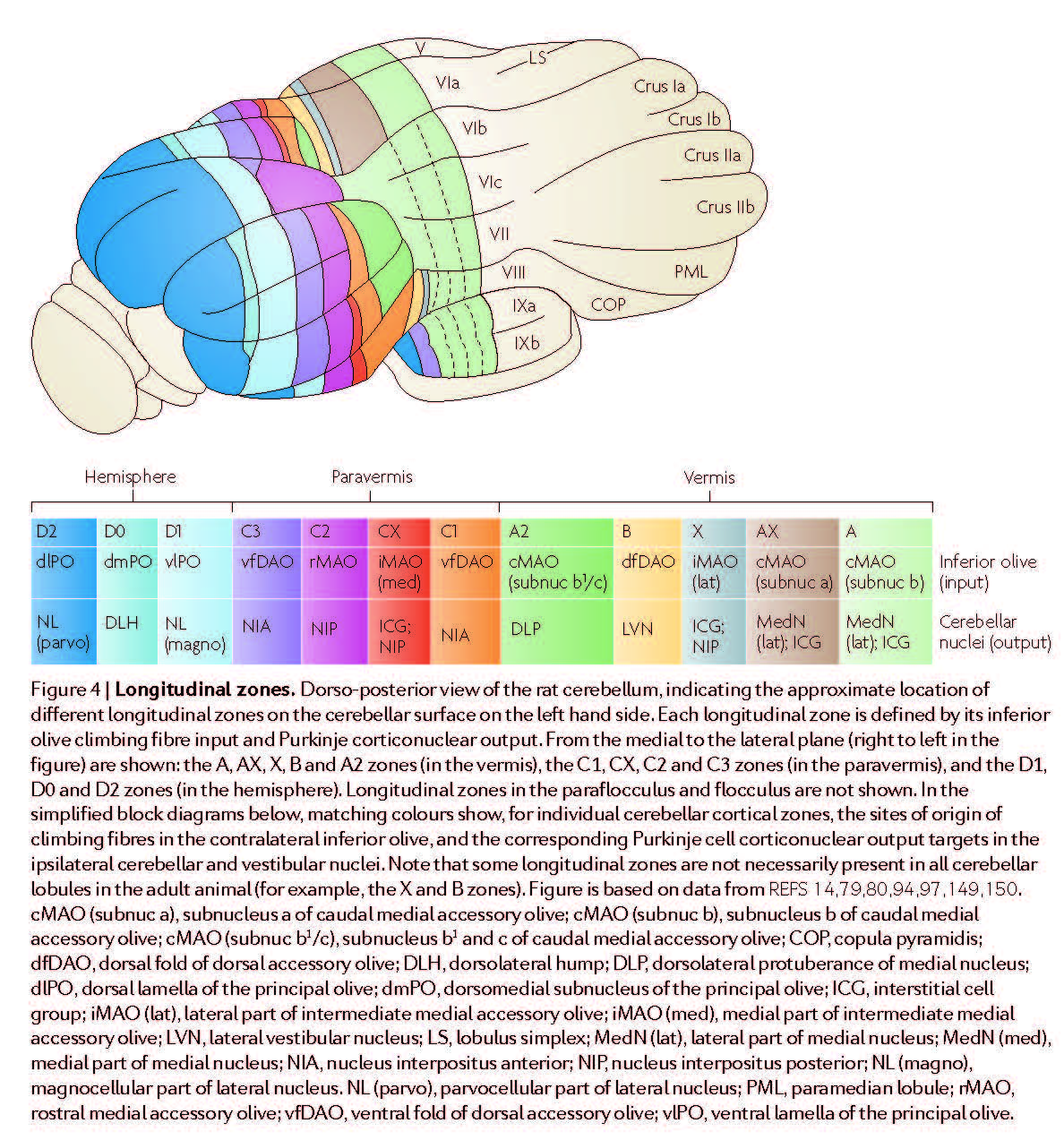
Fig 4 in AppsR+HawkesR-2009 [AppsR+HawkesR-2009].¶
674-675
Modules and microzones.
Anatomical tract tracing has also been used to describe the way in which olivocerebellar (climbing fibre) afferents and corticonuclear (Purkinje cell) efferents are linked to form discrete complexes, the cortical component of each of these ‘modules’ being a longitudinal zone of Purkinje cells2,79. Anatomically defined modules have been extended to include other connections, notably the nucleo-olivary and reciprocal olivo-nuclear projections86. In the same way that Purkinje cell stripes are evolutionarily conserved, cerebellar modules are remarkably similar in a range of species (see Supplementary information S1 (table)). S1 (table) The behavioural significance of these modules is beyond the scope of this Review but their conservation implies that each subserves a similar function in different species (for example, regulation of spinal reflexes and limb movements by the paravermal modules87; and control of compensatory eye movements by modules in the flocculus88).
Some longitudinal zones can be further split into smaller units called ‘microzones’82,89. In particular, highresolution electrophysiological mapping from medial to lateral across the width of the vermal b zone or the paravermal C3 zone shows small groups of Purkinje cells with distinct climbing fibre receptive fields that arise from different body parts. Each Purkinje cell group typically occupies a narrow, rostrocaudally oriented strip of cortex within the broader longitudinal zone (each zone ~1 mm and each microzone ~100–300 μm wide). microzones mapped in the vermal b zone form a regular array of olivo-cortico-nuclear ‘microcomplexes’ that are thought to control different aspects of the motor functions handled by the broader module82. However, the most extensively studied microzones lie within a small part of the C3 longitudinal zone in the paravermis of lobule V in cats, and their organization seems to be rather different89. Whereas individual microzones in the b zone are thought to extend the entire rostrocaudal length of the zone (more than 100 mm in cat)82, microzones in C3 tend to be much shorter, usually extending across no more than a few adjacent cerebellar folia. In addition, microzones located in different parts of the paravermal cortex can have the same climbing fibre receptive field characteristics (for example, there are at least four separate ‘eyeblink’ microzones in each paravermis90). This has led to the concept that spatially separated collections of microzones with common climbing fibre input — termed ‘multizonal microcomplexes’ — may be important for the parallel processing and integration of information from mossy fibre inputs derived from multiple sources3,89.
Some anatomical data are consistent with such a possibility (for example, see REF. 91), but physiological studies to fully test this hypothesis are currently lacking. Anatomical tracer studies have also revealed a correspondingly detailed map within the inferior olive, with subgroups of olivary cells providing climbing fibres to different parts of the same longitudinal zone (for example, see REF. 92 and FIG. 4). In some cases the resolution of the anatomical mapping has been sufficient to reveal this connectivity at a level that might correspond to microzones93,94. Longitudinal zones are therefore most probably composite entities, and the basic operational unit of the cerebellar cortex is narrower, possibly an individual microzone or, in the case of the paravermis, an assembly of microzones forming a multizonal microcomplex3.